содержание .. 111 112 113 114 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 113
вает время, необходимое для решения какой-то пространственной задачи, как
показатель активности соответствующих психических процессов (Posner, 1978).
Например, в одном из классических исследований (Shepard, Metzler, 1971)
испытуемые должны были рассматривать пары изображений трехмерных объектов.
В каждой пробе от испытуемого требовалось указать, одинаковы ли два объекта по
форме или один является зеркальным отображением другого. Два объекта могли
различаться также и по своей ориентации (рис. 26.9). Было показано, что время,
необходимое для того, чтобы определить, одинаковы ли объекты по форме,
закономерно возрастало при увеличении угловых различий между парами
рисунков, предъявляемых испытуемому (рис. 26.10). Из этих наблюдений обычно
делают вывод, что, прежде чем сравнивать по форме оба изображения, испытуемые
поворачивают «в уме» мысленный образ одного объекта, чтобы поставить его в
одинаковое положение с другим. Хотя истинная природа образа, используемого в
этом эксперименте, еще остается спорной (Kosslyn, 1981; Cooper, 1982), наиболее
распространенная прямолинейная интерпретация этих данных состоит в том, что
здесь имеет место какая-то разновидность психического образа и что процессы
вращения и сравнения выполняются один за другим. Тот факт, что время реакции
является функцией от углового смещения предъявляемых изображений, служит
основанием для заключения о том, что мысленное вращение объекта занимает
определенное время (около 30 мс на каждые 20 градусов) (Cooper, Shepard, 1973).
Проверка способности к зрительному распознаванию объектов независимо от их
относительной пространственной ориентации используется в нескольких тестах
для оценки интеллекта и способностей личности (Petrusic et al
., 1978); было бы
интересно сравнить на этой основе представителей разных видов животных (см. разд. 27.1).
Однако до сих пор систематические исследования проведены на животных только одного
вида, и результаты представляются достаточно удивительными.
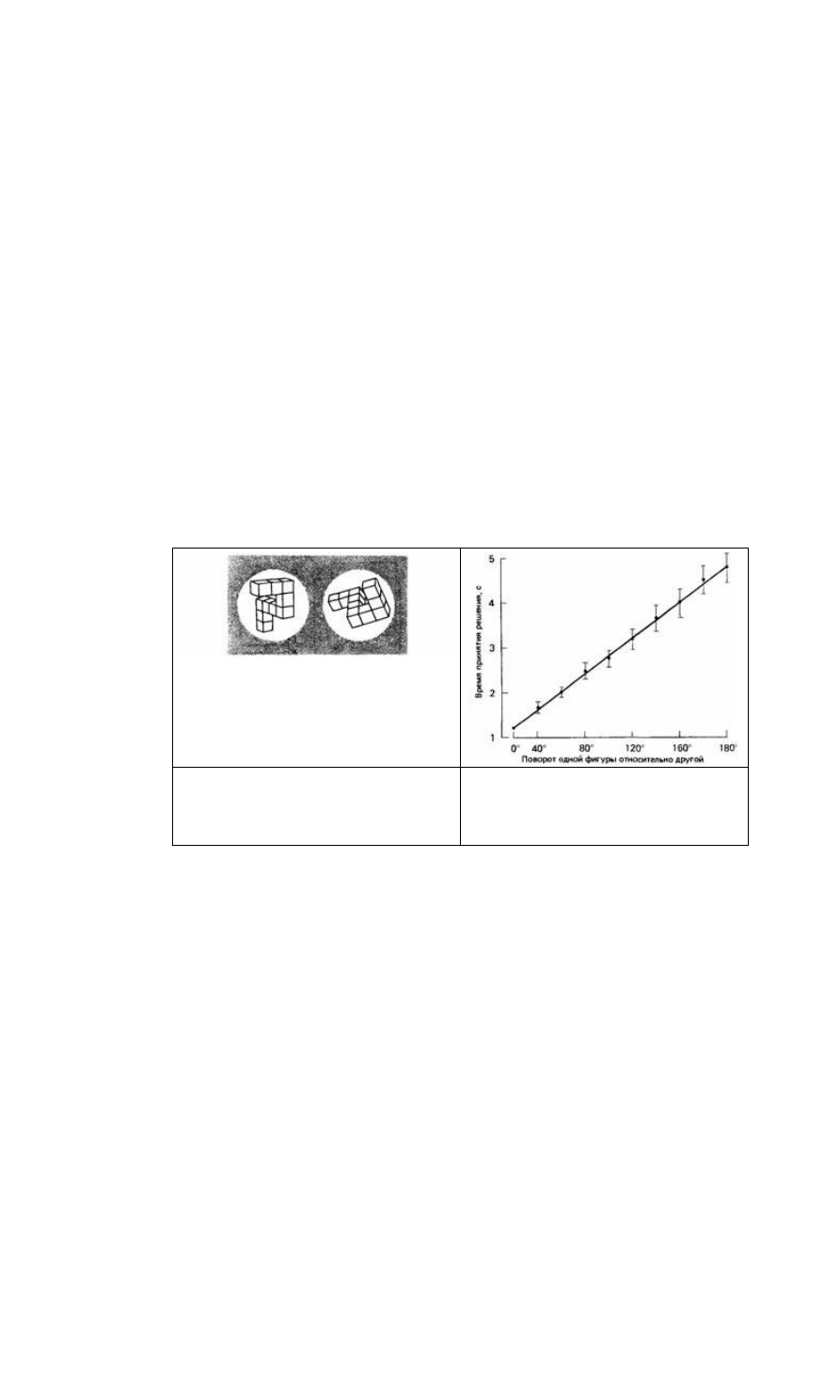
Рис. 26.9. Пример пары фигур, которые
используются для «психологической
хронометрии». Это идентичные фигуры, но как
бы повернутые на 80 одна относительно другой.
(Shepard, Metzler, 1971.)
Рис. 26.10. Время, которое требуется для
установления идентичности двух схожих фигур,
как функция ориентации этих фигур по
отношению друг к другу. (Shepard, Metzler, 1971.)
Валери Холлард и Джуан Делиус (Valerie Hollard, Juan Delius, 1983) обучали
голубей в камере Скиннера различать прямое и зеркальное изображения фигур,
которые предъявлялись птицам ориентированными различным образом (см. рис.
26.11). Затем они измеряли время реакции голубя в тестах на инвариантность по
отношению к повороту. Когда эта часть исследований была завершена, камеру
размонтировали и тестовую панель, на которой находились зрительные сигналы и
клавиши, использовали в аналогичных экспериментах на людях. Таким образом, на
основе одних и тех же конфигураций стимулов можно было непосредственно
сравнить, как справляются с этой задачей голубь и человек.
Результаты показали, что если судить по сделанным ошибкам, то люди и голуби
способны решать эти задачи пример-
453