содержание .. 60 61 62 63 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 62
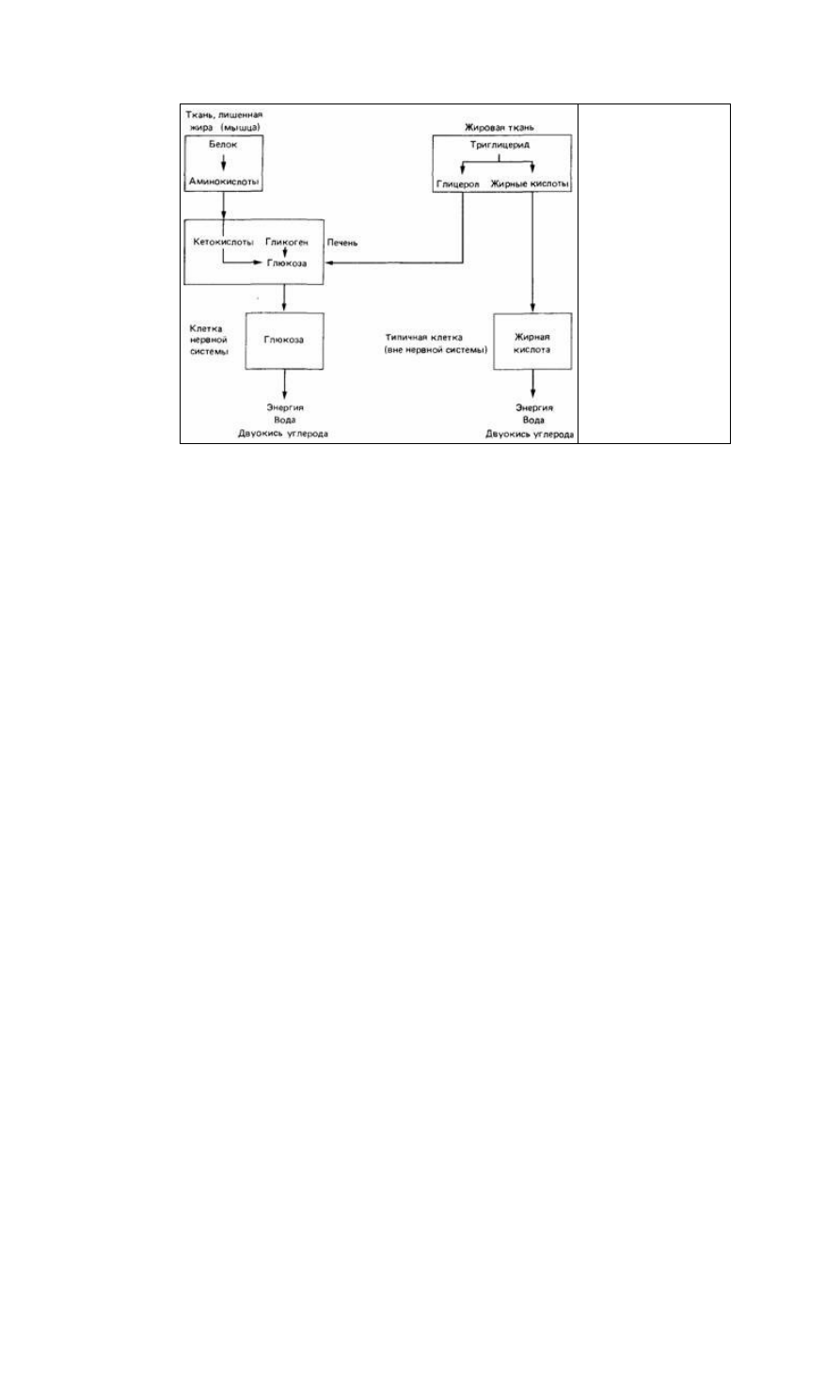
Рис. 15.9. Основные
источники энергии,
доступные во время
голодания (По Toatcs, 1980.)
ных компонентов. Так, липаза разлагает молекулы жира на глицерол и жирные
кислоты, а трипсин и химотрипсин расщепляют специфические аминокислотные
связи в белках. Процесс пищеварения и участвующие в нем ферменты значительно
различаются у разных видов животных. У одних, например у плоского червя
планарии, полный пищеварительный тракт отсутствует, у других, в частности у
травоядных позвоночных, очень сложные пищеварительные системы позволяют
справляться с растительными материалами типа клетчатки, которую другие
животные переваривать не в состоянии. Продукты переваривания переходят в
кровоток отчасти за счет диффузии, а отчасти за счет активного транспорта через
кишечную стенку.
Процесс переваривания и характер питания часто тесно связаны. Многие животные
изменяют потребление пищи в зависимости от питательной ценности продуктов
пищеварения. В регуляции этого типа участвует много механизмов, из которых
самый простой состоит в прямом обнаружении вещества, как это
предположительно происходит с ионами натрия. Высокое и достаточно постоянное
его содержание в жидкостях организма жизненно важно. Натрий участвует во
многих фундаментальных физиологических процессах, включая распространение
нервных импульсов (см. гл. 11). В природе он доступен для животных в виде
хлористого натрия (поваренной соли), но встречается нечасто. Поэтому
неудивительно, что животные должны обладать специальной тягой к натрию. Они
обнаруживают его в пище двумя главными способами. Во-первых, для
большинства позвоночных поваренная соль обладает выраженным вкусом (см.
разд. 12.1). Во-вторых, натрий оказывает сильное действие на жидкую среду
организма, как было указано выше, и его дефицит приводит к секреции гормона
альдостерона из коры надпочечников, который вызывает реабсорбцию натрия из
мочи, образующейся в почках.
Тяга к натрию, по-видимому, является врожденной, но многие животные хорошо
научаются распознавать и запоминать места, где находятся его источники. Так,
например, крысам можно предоставить выбор между пресной и соленой водой в
качестве награды в лабиринте. Крысы, которым не давали пить, научались
оказывать предпочтение тому месту в лабиринте, где была пресная вода. Если тех
249