содержание .. 54 55 56 57 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 56
моторной регуляции у рыб и амфибий. Как было указано в гл. 13. электрическая
стимуляция тектума у жаб вызывает поворот головы, а также схватывание и
проглатывание корма. У эволюционно более продвинутых позвоночных высший
отдел головного мозга в большей степени способен контролировать движения, но
автоматические компоненты локомоции по-прежнему регулируются прежде всего
его стволом и спинным мозгом.
У млекопитающих кортикоспинальный тракт является самым важным проводящим
путем, участвующим в произвольной регуляции движения. Он начинается в
двигательной коре и проходит через средний мозг и мозговой ствол к спинному
мозгу. Эта система, иногда называемая пирамидной, имеется у всех
млекопитающих, за исключением очень примитивных однопроходных (например,
утконоса и австралийской ехидны). У сумчатого кистехвостого кускуса
(Trichosaunis) пирамидные аксоны доходят только до грудного отдела, где они
иннервируют передние конечности. Задние конечности иннервируются
экстрапирамидной системой.
Экстрапирамидная система включает все двигательные пути, не входящие в
кортикоспинальную или пирамидную системы. Она считается более примитивной
системой, чем пирамидная. У животных со слаборазвитой корой больших
полушарий или вовсе лишенных ее базальные ганглии экстрапирамидной системы
являются основными центрами моторного контроля. Они особенно хорошо развиты
у птиц, у которых практически нет коры, но стриатум крупнее, чем у
млекопитающих. Таким образом, в ходе эволюции кора больших полушарий, по-
видимому, стала вторым источником двигательной координации, действующим
через пирамидную систему.
У обезьян нейроны в разных частях кортикоспинальных путей меняют характер
своей импульсации при произвольных • движениях глаз, передних и задних
конечностей (Evarts. 1968). Искусственная стимуляция двигательной коры
вызывает реакции отдельных мышц или отдельных моторных единиц в мышце.
Более обширная стимуляция приводит к отдельным
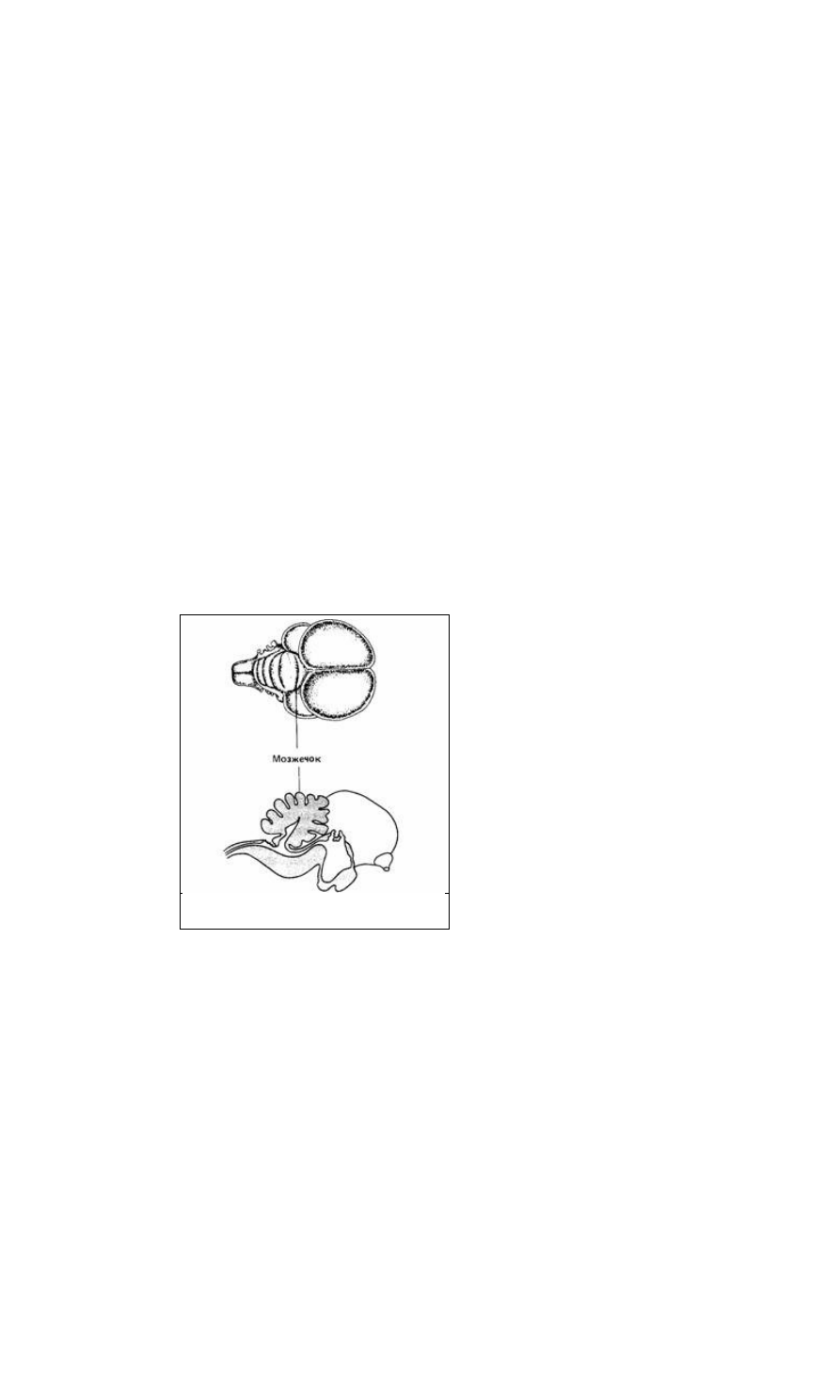
Рис. 14.4. Головной мозг голубя с крупным
мозжечком
движениям целой конечности. Можно нанести карту на поверхность коры в
соответствии с теми частями тела, которые движутся в ответ на ее электрическую
стимуляцию. Можно построить такую же карту для сенсорной коры, на которой
разные части тела представлены по-разному в соответствии с их сенсорным
значением для данного вида. Произвольные выученные движения вызываются
корковыми пирамидными нейронами, а рефлекторное поддержание двигательных
реакций и позы управляются расположенными поблизости экстрапирамидными
нейронами. Число пирамидных нейронов у человека оценивается в один миллион
(Prosser, 1973).
Другой важной для координации частью головного мозга позвоночных является
мозжечок (рис. 14.4). Он участвует в позных рефлексах или в управлении
движением косвенно, являясь контролером и координатором нервной активности
при ориентации, сохранении равновесия и других тонких реакциях, связанных с
регуляцией движения. Основная нейронная организация мозжечка одинакова во
всех классах позвоночных - он претерпел меньше эволюционных изменений, чем
любая другая часть головного мозга. Мозжечок получает информацию от органов
зрения,
225