содержание .. 52 53 54 55 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 54
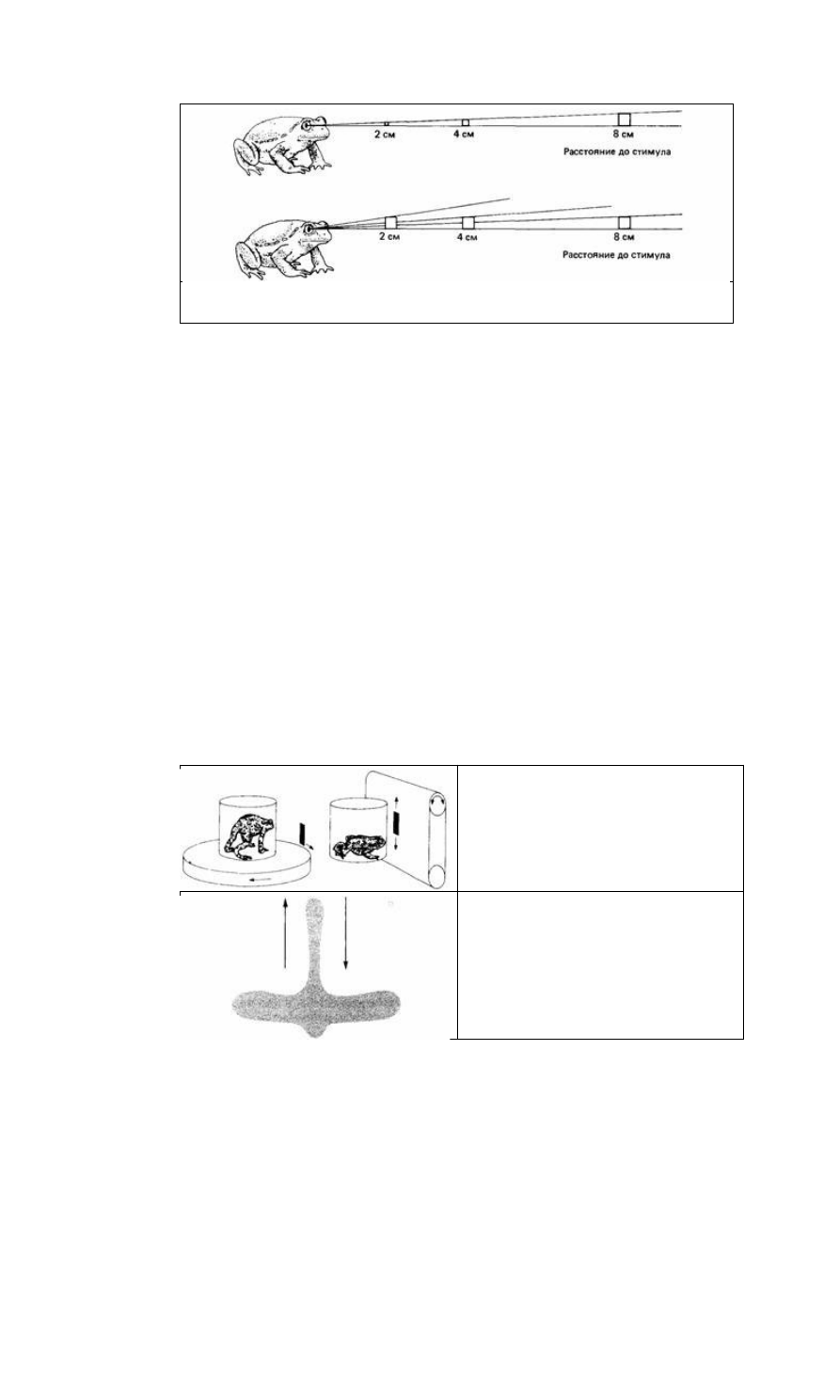
Рис. 13.7. Молодые жабы (вверху) оценивают размеры по углу зрения, а взрослые жабы (внизу)
могут судить о размерах независимо от угла зрения. (По Ewert, 1980.)
двумерными фигурами Робинсон обнаружил, что двусторонняя симметрия,
вероятно, служит одним из признаков добычи, на который обычно ориентированы
хищники.
Те же принципы применимы и к узнаванию жертвами хищников. Так, например,
силуэт ястреба, движимый над утятами или гусятами, вызывает реакцию страха,
когда движется только в определенном направлении (рис. 13.9). Это объясняется
тем, что короткая шея и длинный хвост характерны для ястреба, а длинная шея и
короткий хвост - признаки летящего гуся (Tinbergen, 1951). Жабы избегают фигур,
похожих на змею с поднятой головой (рис. 13.10). Пиявка, движущаяся толчками,
воспринимается как добыча, если ее передняя присоска находится на почве, но если
эта присоска поднята в воздух, жаба принимает пиявку за врага (Ewert, 1980).
Как мы видим, поведенческие исследования (см. обзор Ewert, 1980) показывают,
что жабы хватают мелкие продолговатые объекты, расположенные горизонтально,
но не реагируют на такие же объекты в вертикальном положении (рис. 13.8). Жаба
обычно питается насекомыми, личинками, червяками и т. п. Ее хищническое
поведение состоит из ориентации головы и туловища, зрительной фиксации
добычи, захватывания ее в результате вытягивания шеи и языка, глотания и
вытирания морды передними лапами (рис. 13.11). Чтобы вызвать хищническое
поведение, нужен мелкий движущийся объект. На крупные движущиеся объекты
жаба отвечает оборонительным поведением. Хотя для изучения сенсорных
процессов могут быть применены разные чисто поведенческие методы,
физиологическое исследование тоже может давать ценные сведения о деятельности
органов чувств и о типе информации, которую они посылают в мозг. Но для того
чтобы установить, как мозг использует такую информацию, тре-
Рис. 13.8. Жабы реагируют положительно на
прямоугольники, движущиеся вперед узкой
стороной, и отрицательно на движущиеся вперед
широкой стороной. (По Ewert, 1980.)
Рис. 13.9. Этот силуэт похож на ястреба, если его
двигать в одном направлении, и на гуся, если его
двигать в противоположном направлении.
217