содержание .. 55 56 57 58 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 57
тает взад и вперед, не продвигаясь против ветра (т. е. под прямым углом к нему).
Когда ветер приносит запах, угол полета увеличивается и животное движется
зигзагами против ветра. Изменения направления связаны с границами запахового
следа (рис. 14.9). При снижении концентрации запаха ниже определенного уровня,
в частности у края его струи, животное начинает двигаться в противоположном
направлении. Эти повороты не связаны с направлением ветра, а зависят от
внутреннего эталона, или идиотетической информации. Таким образом в поисках
полового партнера летящая бабочка использует сочетание зрительных,
анемотаксических (связанных с ветром) и идеотетических ориентационных
механизмов.
14.3. Принцип реафферентации
Высокоразвитая система ориентации должна быть способна отличать стимулы из
внешнего мира от стимуляции, создаваемой самим животным. Так, например, что
касается зрения, то у человека перемещение объектов во внешнем мире вызывает
движение изображения по сетчатке, которое мы воспринимаем. Однако
произвольное движение глаз тоже вызывает перемещение изображения на сетчатке,
но оно уже не воспринимается. Каким-то способом мозг отличает перемещение
изображения по сетчатке, независимое от животного, от перемещения,
вызываемого движением самого глазного яблока.
Для объяснения этого явления предложены две теории - оттока и притока. Согласно
теории оттока, основы которой заложил Гельмгольц (Helmholtz, 1867). команды
глазным мышцам о движении глазного яблока сопровождаются параллельными
сигналами, идущими к компаратору в головном мозгу. Здесь они сравниваются с
приходящими зрительными сигналами (рис. 14.10). Согласно теории притока,
созданной Шеррингтоном (Sherrington, 1918), рецепторы наружных глазных мышц
посылают сообщения в мозговой компаратор при любых движениях глаз (рис.
14.10). В обеих теориях компаратор оценивает оба приходящих
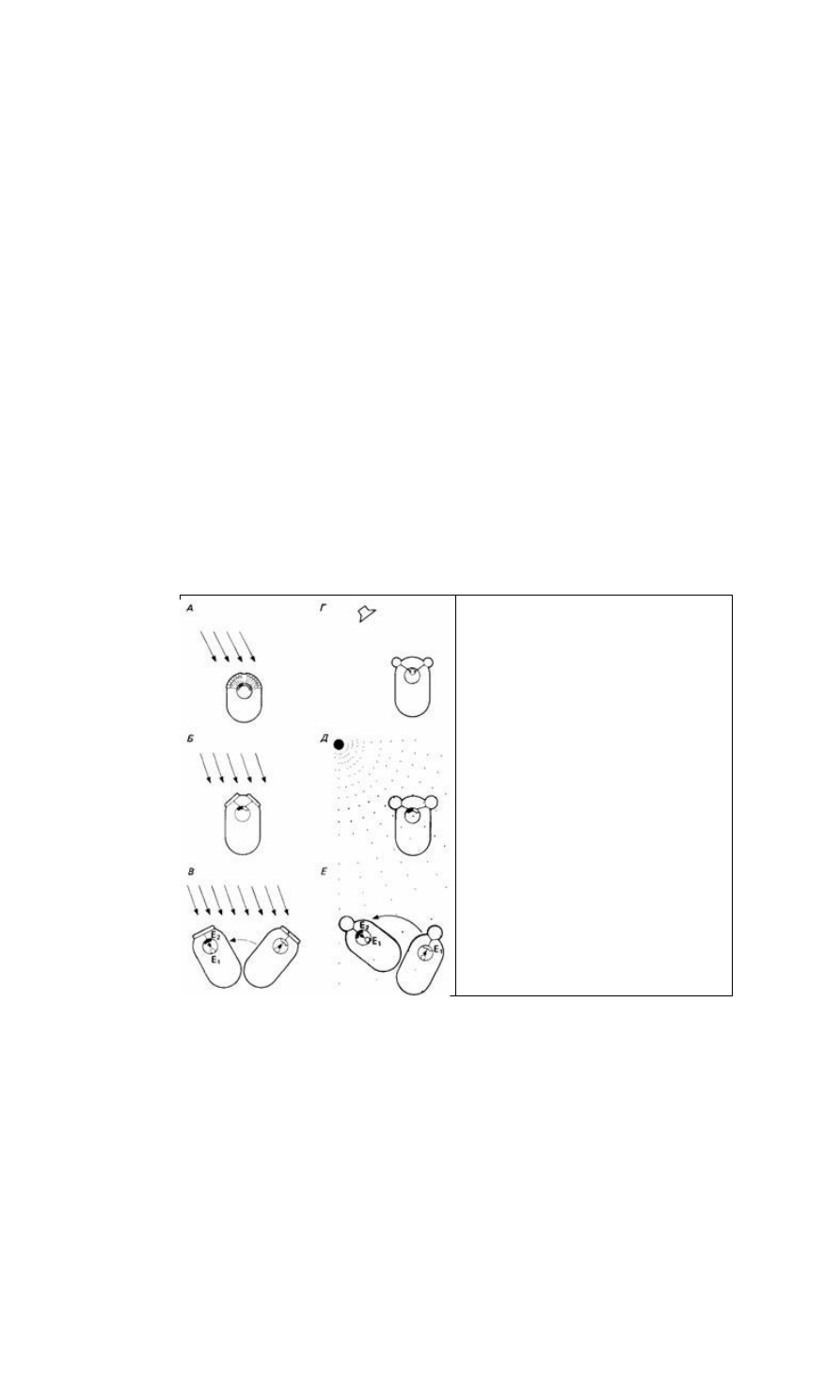
Рис. 14.8. Схематическое изображение некоторых
основных принципов сенсорной ориентации. А.
Направление стимуляции (например, снега)
регистрируется растром сенсорных рецепторов. Б.
Направление регистрируется посредством
сравнения одновременной стимуляции двух
рецепторов. В. Имеется только один рецептор, и
животное проводит последовательные сравнения,
передвигая туловище. Г. Время прихода стимула
(например, "звуковых волн) сравнивается двумя
рецепторами. Д. Градиент стимуляции (например,
химической) peгистрируется посредством
сравнения данных, получаемых двумя рецепторами.
Е. Градиент регистрируется одним рецептором,
когда животное движется, обследуя разные точки
пространства. (Из The Oxford Companion to Animal
Behaviour, 1981.)
сигнала и определяет, соответствуют ли зрительные сигналы тому движению,
которое следовало ожидать на основании второго сигнала. Если сигналы не
соответствуют друг другу, значит, какая-то часть движения должна была быть
вызвана внешними причинами.
Наружные глазные мышцы содержат
229