содержание .. 47 48 49 50 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 49
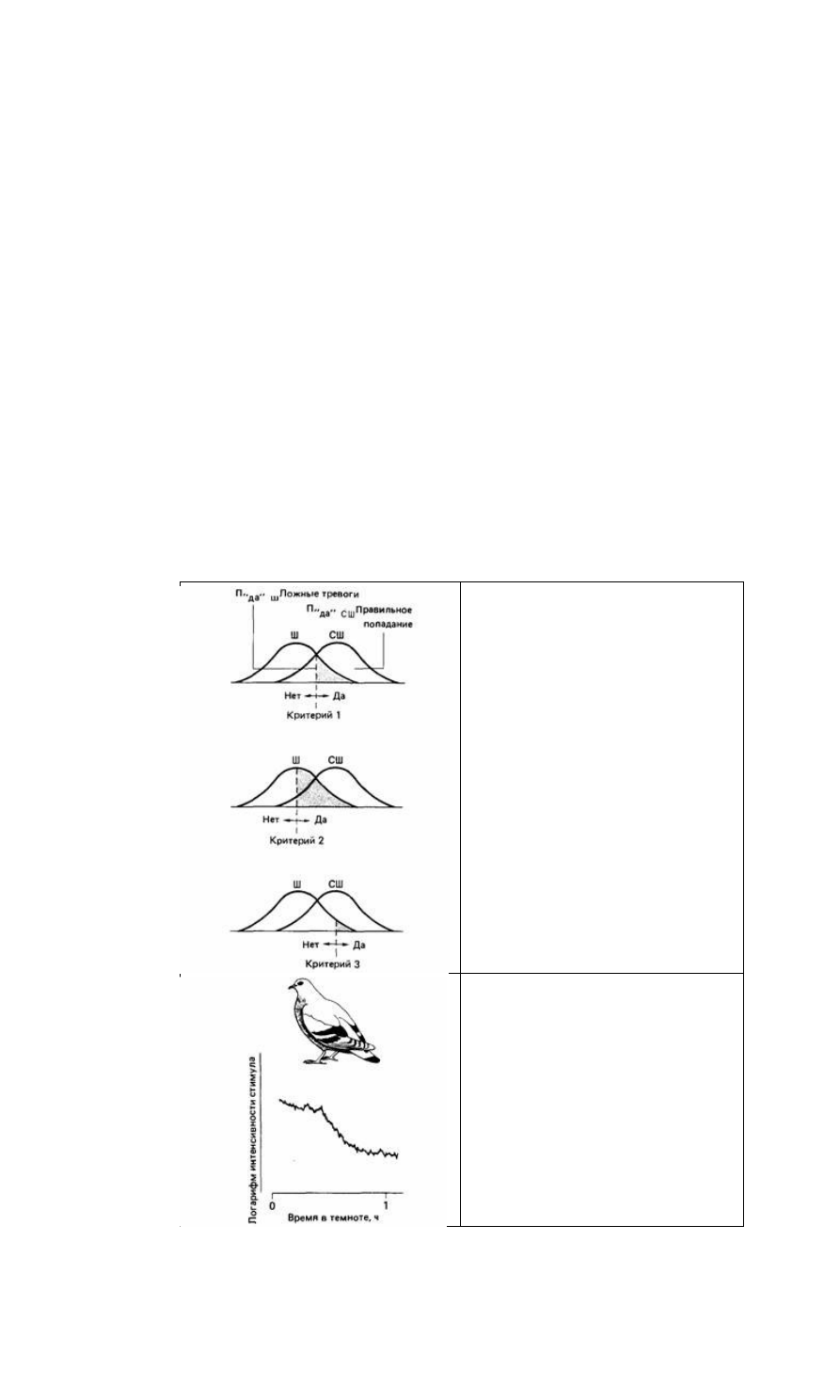
как ложных тревог, так и правильных попаданий, и наоборот (рис. 12.18). При
усилении стимула число ложных тревог снижается, а число попаданий растет.
Психофизическими методами изучались многие стороны поведения животных.
Первым исследованием такого рода, по-видимому, была работа Блафа (Blough,
1955), который изучал темновую адаптацию у голубей. Этот автор научил голубей
клевать два ключа, расположенных непосредственно под освещенным окошечком.
Свет проникал в аппарат только от этого источника. Птицы были научены клевать
ключ А, когда видели стимул, и ключ Б, когда не видели его. Клевание ключа А
автоматически приводило к потускнению стимула, а клевание ключа Б повышало
его яркость. Обученную птицу тестировали следующим образом.
Сначала стимульное окошечко ярко освещено, и птица клюет только ключ А.
Каждый удар клювом снижает яркость стимула, но птица продолжает клевать этот
ключ, пока стимул не станет таким слабым, что будет ниже абсолютного порога
для голубя. В этот момент птица начинает клевать ключ Б, из-за чего стимул
становится ярче. Когда он снова становится видимым птице, она возвращается к
ключу А. В течение часа птица попеременно клюет ключи А и Б, и яркость стимула
колеблется в области абсолютного порога птицы. Эта процедура позволяет
проследить кривую темновой адаптации голубя. Сначала птица адаптирована к
свету, но по мере потускнения стимула начинает адаптироваться к темноте. Иными
словами, ее порог постепенно меняется так, как показано на рис. 12.19. Обратите
внимание на характерный перегиб этой кривой, соответствующий переходу от
колбочкового к палочковому зрению (см. рис. 12.14).
12.5. Фильтрация стимула
Анатомические и физиологические исследования органов чувств и связанных с
ними отделов нервной системы могут дать ценную информацию о сенсорных
способностях животного. Однако сами по себе они не позволяют сделать оконча-
Рис. 12.18. Гипотетические эффекты разных
критериев решения согласно теории обнаружения
сигнала. Вероятность ответов «да» соответствует
области справа от критерия, а вероятность ложных
тревог темным областям. Предполагается, что стимул
и шум остаются поименными.
Рис. 12.19. Кривая темновой адаптации голубя,
полученная психофизическим методом. (По Blough,
1955.)
197