содержание .. 40 41 42 43 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 42
гой группы, в том числе у позвоночных (Bullock, 1977). В табл. 11.1 приведены эти
основные типы. Тела сенсорных нейронов в таких рецепторах лежат близ
чувствительной поверхности и не сгруппированы в сенсорные ганглии. Как
показано на рис. 11.15, у одних членистоногих сенсорных нейронов мало, у других
- много.
Ракообразным свойственно много типов механорецепторов, в том числе
статоцисты. Особыми внутренними механорецепторами являются хордотоналъные
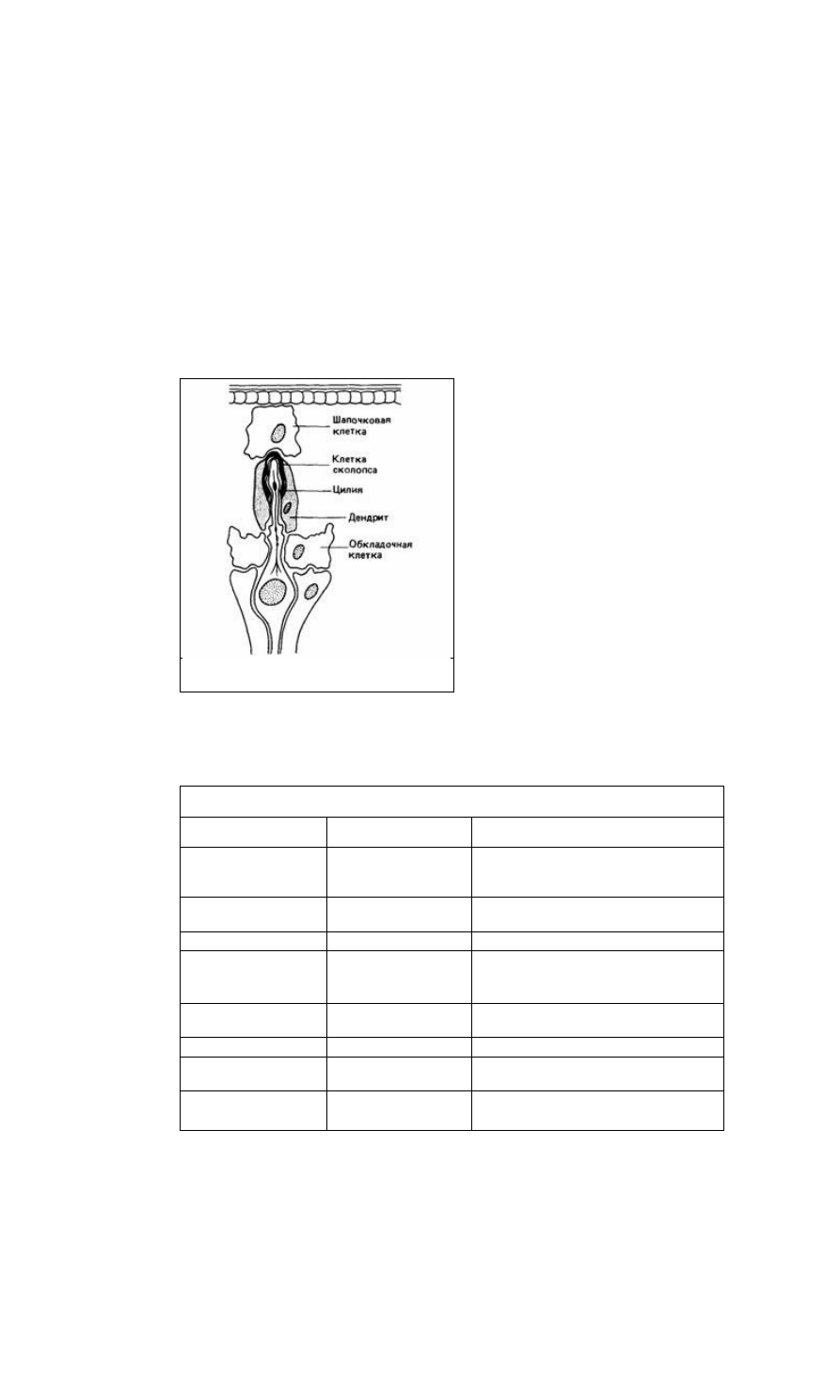
органы, которые обычно выполняют функции проприоцепторов, отвечающих на
механическое смещение (рис. 11.16). Они могут также участвовать в определении
направления ветра, скорости течения воды, скорости полета, направления силы
тяжести и разных типов вибраций. Механорецепторы обнаружены также в связках
и мышцах, откуда они посылают информацию о положении конечностей и
движении.
Особый интерес представляют фоторецепторы членистоногих, один из которых
показан на рис. 11.17. Вообще членистоногие отличаются сложностью нервной
системы и многими направлениями ее эволюционного развития, в том числе по-
Рис. 11.16. Хордотональный механорецептор
насекомого. (По Gray, 1950.)
степенным переходом от сегментарного строения, свойственного кольчатым
червям, к крупному мозгу, образованному многими слившимися ганглиями. Они
обладают удивительно богатым поведенческим репертуаром. Некоторые виды,
например медоносная пчела (подробно описанная в гл. 23), по сложности
поведения
Таблица 11.1. Некоторые типы кутикулярных рецепторов членистоногих
Структура
Морфологическое строение Функция (известная или предполагаемая)
Трихоидная сенсилла
Сенсорные волоски и
щетинки
Механорецепторы, проприоцепторы, звук,
контактная хеморецепция, влажность,
обонятельная в разных местах
Хетоидная сенсилла
Сенсорные шипики и
щетинки
Механорецепторы, проприоцепторы
Лопастевидная сенсилла Сенсорные чешуйки
Механорецепторы
Базиконическая сенсилла Короткие толстые
волоски; малое или
большое число нейронов
Механорецепторы, контактные
хеморецепторы
Целоконическая сенсилла Погруженные
кутикулярные конусы
Обонятельные рецепторы или рецепторы
влажности
Ампуловидная сенсилла Сенсорные трубочки
Обонятельные рецепторы
Колоколовидная
сенсилла
Кутикулярные бугорки
Дирекциональные измерители натяжения
Плакоидная сенсилла
Кутикулярные и пористые
пластинки
Неизвестна; во множестве на антеннах пчел
169