содержание .. 19 20 21 22 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 21
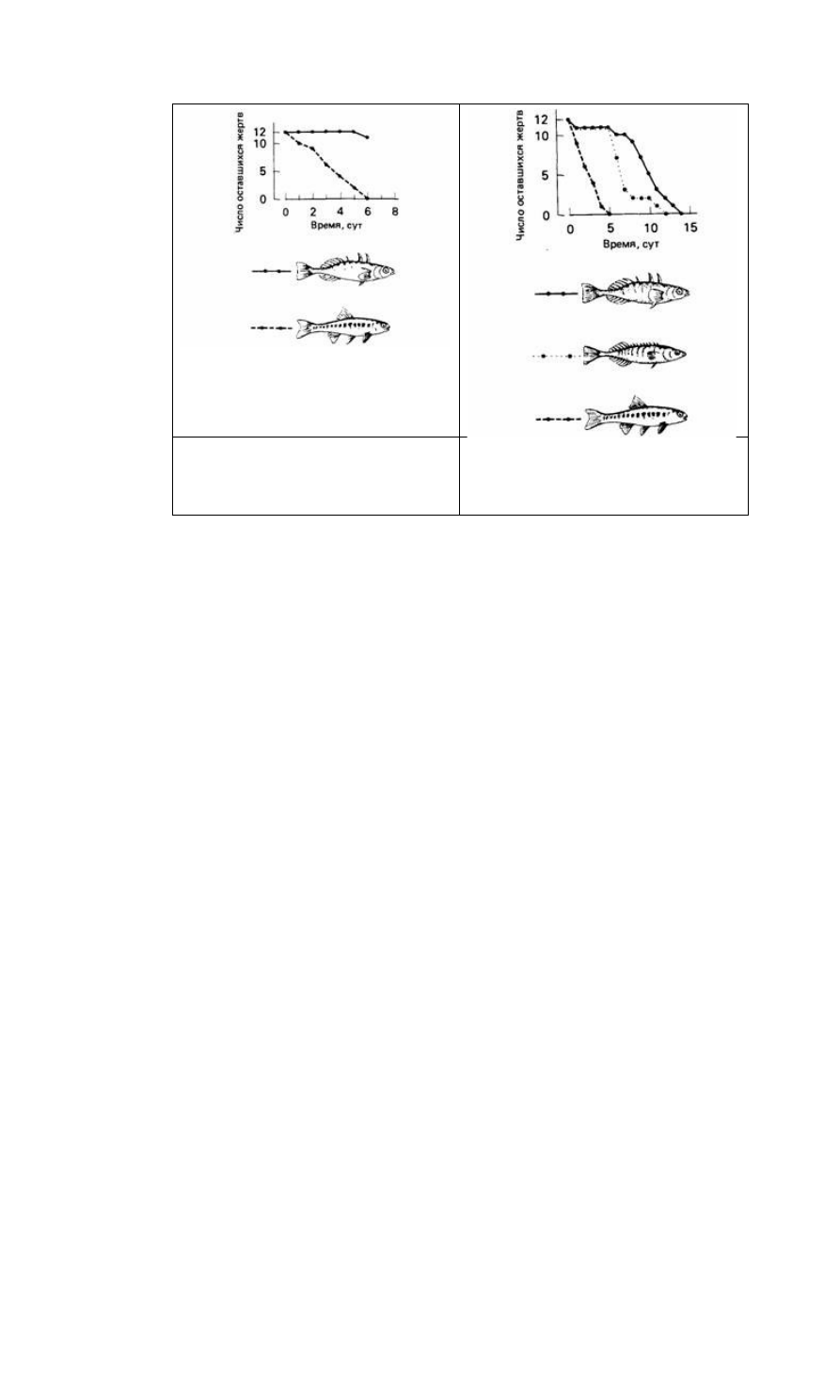
Рис. 6.4. Влияние хищничества щуки на
смешанную стайку из 20 трехиглых колюшек
(сплошная линия) и 20 гольянов (штриховая
линия). (По Hoogland и др., 1957.)
Рис. 6.5. Влияние хищничества щуки на
смешанную стайку из 20 трехиглых колюшек
(сплошная линия). 20 десятииглых колюшек
(пунктирная линия) и 20 гольянов (штриховая
линия). (По Hoogland и др., 1957.)
гнезда, но начинали воздушные атаки на хищника. Они не улетали от других чаек,
ежей или ворон, а сами нападали на них, если те подходили близко к гнезду.
Эти различия в реакциях на хищников в высшей степени адаптивны. Если чайка
покидает гнездо, она оставляет яйца или птенцов только под защитой окраски, что
не имеет значения, когда нападет сапсан, который опасен только взрослым птицам.
Чайки остаются на гнезде и защищают потомство от ежей и хорьков, которые
опасны только для яиц и птенцов. Энергично атакуя этих хищников, чайки не
только отпугивают их от колонии в данном конкретном случае, но и удерживают от
посещения колонии в будущем. Круук (Kruuk, 1964) обнаружил, что вороны
избегают колонии, а ежи не оставляют попыток проникнуть туда, особенно ночью.
При встрече с лисицей или человеком чайки сталкиваются с такой проблемой:
бросить гнездо - значит подвергнуть опасности потомство, а защищать гнездо -
самим подвергаться опасности. В ситуации такого типа нужно тонкое равновесие
между издержками и выгодами, и чайки, по-видимому, находят в этих случаях
приемлемый компромисс.
Кроме прямой реакции на хищников чайки могут защищать себя различными
непрямыми способами. Паттерсон (Patterson, 1965) показал, что выбор места для
гнезда и время откладки яиц являются важными факторами, влияющими на успех
размножения. Паттерсон заметил, что большая часть обыкновенных чаек на
исследуемой им территории образовывала очень плотные колонии, в которых
гнезда располагались примерно в метре друг от друга. Однако некоторые чайки
гнездятся более чем в ста метрах от колонии. Он показал, что есть заметная разница
в успехе размножения птиц, которые гнездятся внутри колонии и вне ее. Очевидно,
должны быть какие-то преимущества при гнездовании внутри колонии. Чтобы
исследовать их, Паттерсон проследил развитие почти 800 гнезд и обнаружил, что у
чаек, гнездящихся в центре колонии, успех размножения выше, чем у чаек,
гнездящихся ближе к краю, если судить об этом успехе по числу выкормленных
птенцов.
85