содержание .. 10 11 12 13 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 12
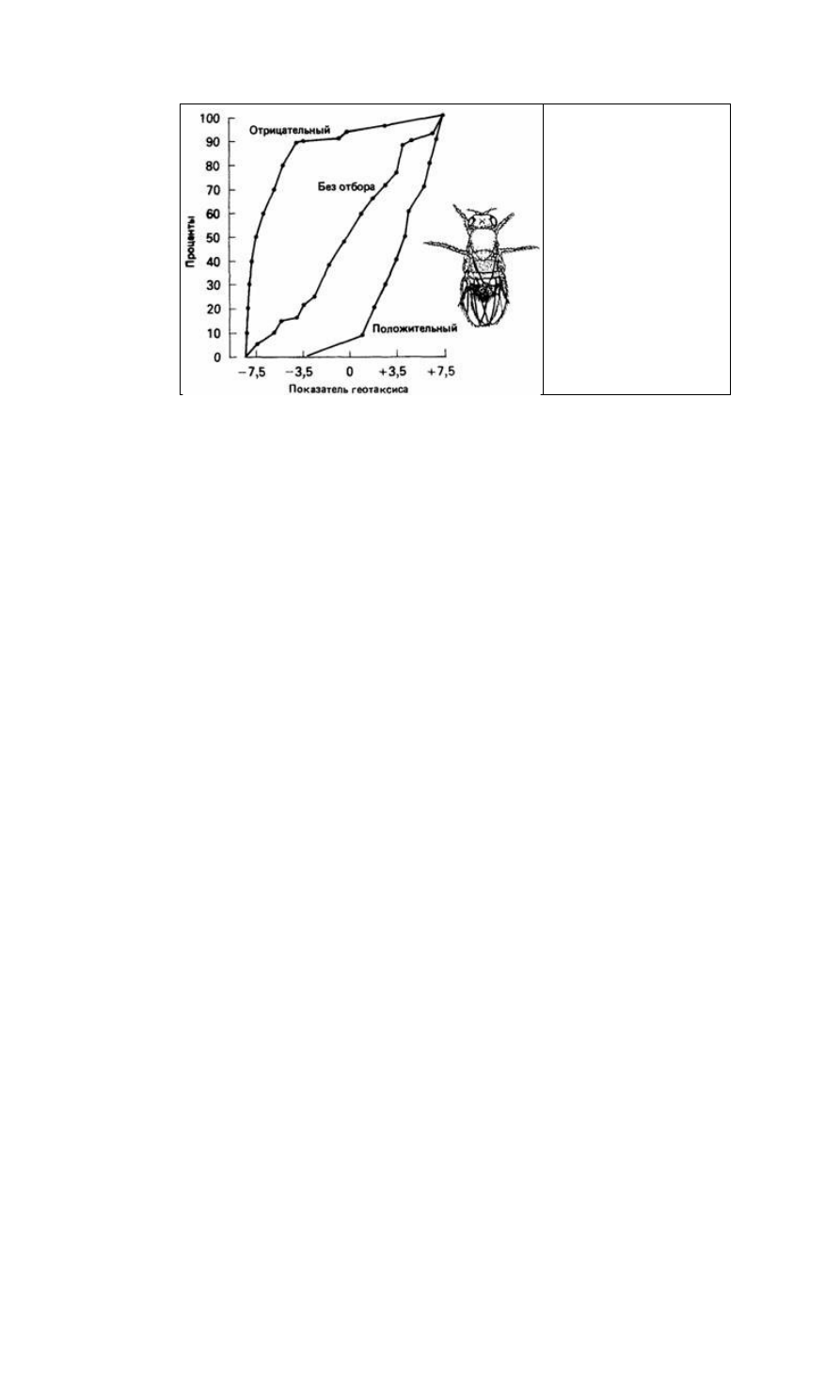
Рис. 4.3. Показатели, полученные
при изучении геотаксиса, у трех
линий Drosophila в лабиринте,
изображенном на рис. 4.2. (По
Hirsch, 1963.)
ли с особой линией, в которой были различные хромосомные инверсии и гены-
маркеры. У Drosophila melanogaster имеются четыре пары хромосом, три пары
больших и одна - маленьких. Гены-маркеры использовались для идентификации
трех больших хромосом. Это были доминантные гены, контролирующие признаки,
отчетливо проявляющиеся в фенотипе. Специальная схема скрещивания позволила
получить самок, которые были либо гомозиготны, либо гетерозиготны по
хромосомам, подлежащим исследованию.
Эти три хромосомы можно было идентифицировать с помощью генов-маркеров
следующим образом: хромосома Х-ген узких глаз (Bar), хромосома II-ген
волнистых крыльев (Cy) и хромосома III - ген, обусловливающий короткие
щетинки (Sb). Тестируемую самку, несущую эти маркеры, скрещивали с самцами
одной из исследуемых линий. Для использования в последующих экспериментах из
потомства отбирали только тех, которые несли все три гена-маркера. При
возвратных скрещиваниях, используя самцов из исходной популяции, получили
восемь возможных генотипов. Следовательно, каждая из трех больших хромосом
гетерозиготна или гомозиготна по S-хромосоме, полученной от тестируемой линии
(s), к которой принадлежал отец. На генотипах этих восьми классов изучали
индивидуальные эффекты и взаимодействие хромосом.
Хирш и Линда Эрленмейер-Кимлинг (Hirsch, Erlenmeyer-Kimling, 1962)
обнаружили, что в популяции, где отбор не проводился, хромосомы X и II
оказывают большое влияние на развитие положительного геотаксиса, а хромосома
III-на развитие отрицательного геотаксиса. В линии, где проводился отбор на
положительный геотаксис, были обнаружены небольшие изменения в хромосомах
X и II, но хромосома III в этом случае оказывала влияние на развитие
положительного геотаксиса. В линии, которая была отобрана на отрицательный
геотаксис, отрицательное влияние хромосомы III увеличивалось, тогда как
положительное действие хромосом X и II было меньше. Эффект от совместного
действия всех трех хромосом был больше в случае отрицательного геотаксиса. Это
неудивительно, так как общая реакция на отбор (см. рис. 4.3) сильнее в случае
отрицательного геотаксиса. Эти результаты показывают, что поведение, связанное
с геотаксисом, контролируется рядом генов, которые находятся во всех трех
больших хромосомах.
Хромосомный анализ широко применяется при изучении различных типов
поведения дрозофилы, в том числе скорости спаривания и других аспектов
ухаживания
49