содержание .. 106 107 108 109 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 108
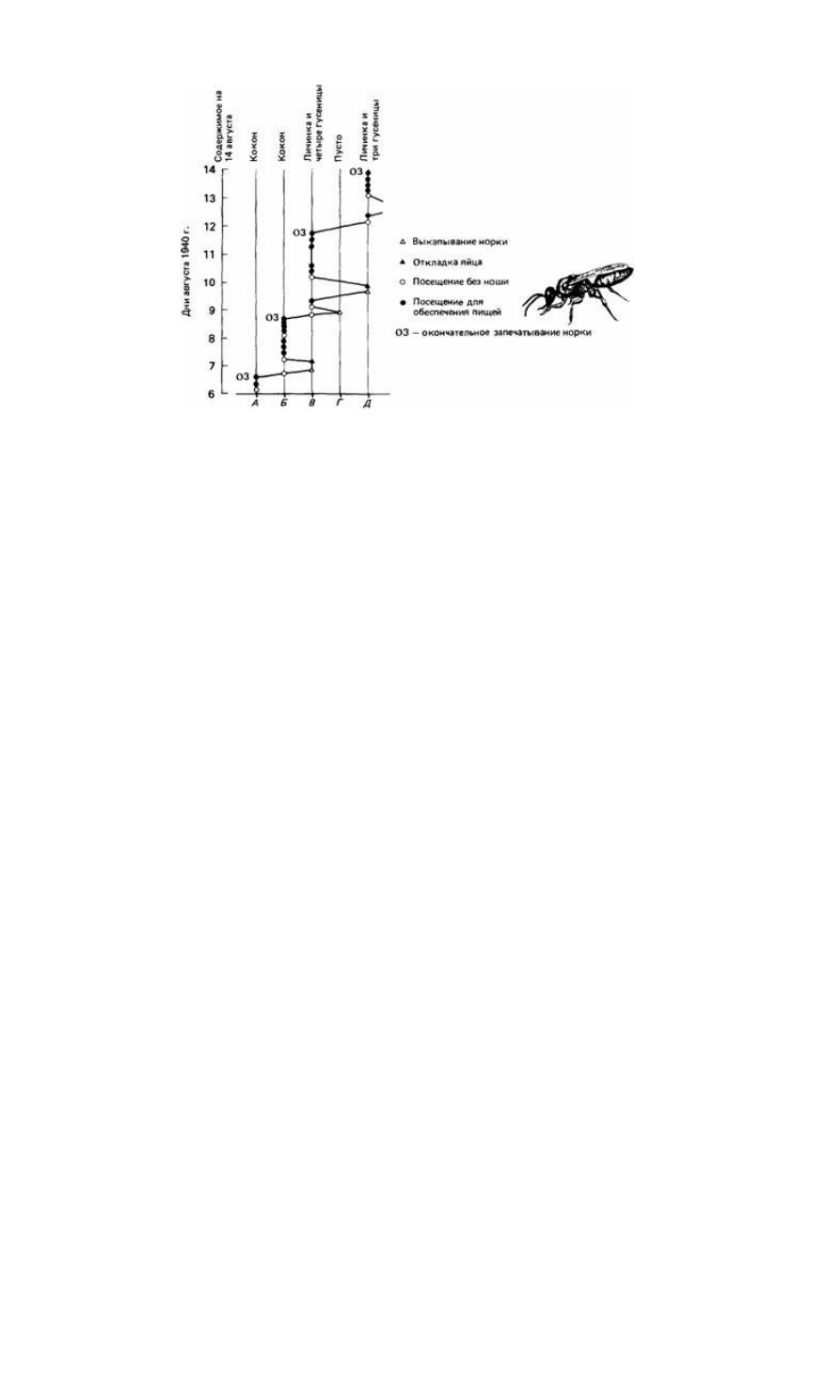
Рис. 25.16. Диаграмма гнездового поведения роющей осы (Ammophila). (По Baerends, 1941.)
сить в гнездо гусениц, если их систематически каждый раз удалять из гнезда, как
только оса их принесет. Этот пример показывает, что сложное поведение может
быть запрограммировано в виде набора жестких правил. Оса ведет себя наподобие
автомата, хотя у нее и есть некоторые стандартные программы поведения,
позволяющие ей выходить из затруднительного положения, например удалять из
норки какие-то препятствия.
Как мы уже видели ранее, прерывание поведения животного при определенных
обстоятельствах маскирует поведение, которое должно было бы проявиться, если
бы не было этого перерыва. Такая ситуация разделения времени предполагает, что
животное следует определенным правилам, которые определяют организацию и
приоритетность поведенческих актов в общей картине поведения. Рассмотрим
конкретный пример. Когда голодная горлица (Streptopelia) ест, либо выбирая зерна
из кучи, либо получая пищу в камере Скиннера, в ее поведении можно наблюдать
типичные паузы длительностью в несколько минут (см. рис. 25.11). Что будет
делать горлица во время этих пауз, зависит от обстоятельств. Если птица имеет
доступ к воде, она будет пить. В противном случае она может чистить перья или
просто неподвижно стоять. В экспериментальных условиях было показано, что на
время возникновения этих пауз никакого влияния не оказывает манипулирование
мотивационными факторами вторичного приоритета, например изменением уровня
жажды. В одном из экспериментов голодным горлицам к каждому крылу
прикрепили по канцелярской скрепке. Во время еды горлицы не обращали на
скрепки никакого внимания, тогда как во время пауз они пытались от них
отделаться. Однако наличие канцелярских скрепок не оказывало влияния на
характер пищевого поведения и не меняло временного распределения пауз
(McFarland, 1970b). Создается впечатление, что в пищевом поведении горлицы как
бы запрограммированы на строго определенное время паузы и что правила,
которые регулируют пищевое поведение горлиц, не находятся под каким бы то ни
было влиянием со стороны других мотивационных факторов, например жажды или
стремления чистить перья, если только эти тенденции не станут сильнее, чем
пищевая. Это типичный случай явления разделения времени.
433