содержание .. 57 58 59 60 ..
ПОВЕДЕНИЕ ЖИВОТНЫХ. Психобиология, этология и эволюция (Д. Мак-Фарленд) - часть 59
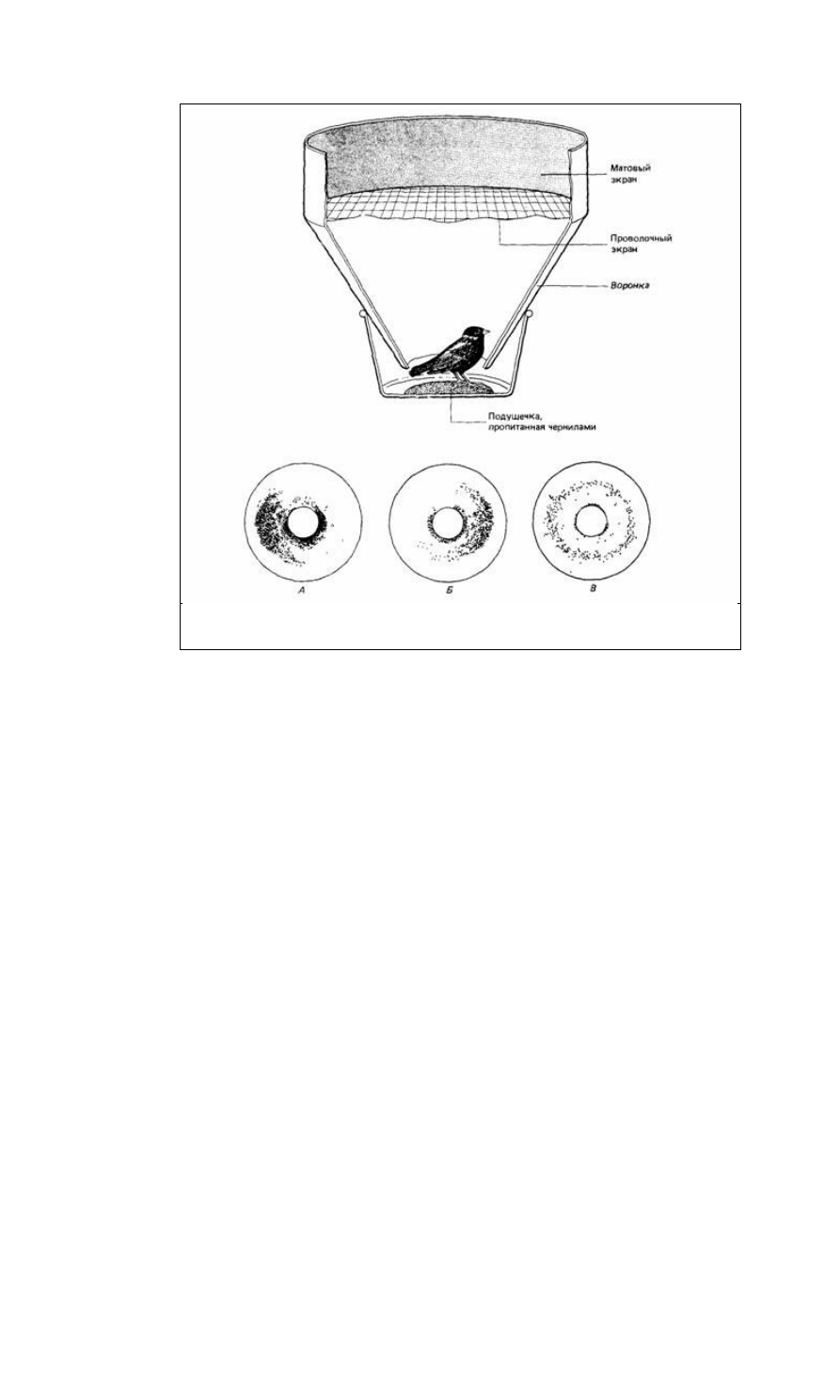
Рис. 14.16. Экспериментальная клетка для измерения миграционного беспокойства. Пытаясь покинуть
клетку, птица оставляет чернильные следы на промокательной бумаге, выстилающей воронку. Внизу -
несколько примеров регистрации. (По Schmidt-Koenig, 1979.)
Было обнаружено (von Frisch, 1967), что ориентация пчел, выражаемая их танцем
(см. гл. 23), зависит от положения солнца, даже когда оно скрыто тучами.
Оказывается, пчелам достаточно видеть только небольшую часть неба, причем
нужная информация соответствует ультрафиолетовой части спектра. При
пропускании ультрафиолетового света солнца через поляризующий фильтр
ориентация танца пчелы менялась в соответствии с углом поляризации.
Опыты с условными сердечными рефлексами (см. рис. 14.14) ясно показывают, что
голуби способны воспринимать вращение плоскости поляризации света (Kreithen,
Keeton, 1974; Delius et al., 1976), но неизвестно, как они интерпретируют эту
информацию. Хотя характер поляризации солнечного света может дать сведения о
местоположении солнца даже тогда, когда небо покрыто тучами, по всей
вероятности, голуби не пользуются своим солнечным компасом при таких условиях
(Schmidt-Koenig, 1979).
237